


Ispirato liberamente, tradotto liberamente, e rielaborato dai lavori di Frank Turek (Stealing from God cap.3), Stephen C. Meyer (Darwin’s Doubt cap.10), Don Batten, Royal Truman, Warner Wallace, Casey Luskin.
Nella Parte 1 abbiamo visto e criticato la teoria evoluzionistica analizzando le mutazioni e un problema statistico. In questa sezione criticheremo il darwinismo analizzando la complessità irriducibile, qualche altro problema statistico e l’inefficienza della selezione naturale. Premettiamo che le categorie trattate in questo articolo sono indipendenti le une dalle altre.
LA SELEZIONE NATURALE E’ UN PROCESSO CHE CONSERVA
Darwin credette che la selezione naturale potesse selezionare persino la variazione più piccola, ma in realtà, come spiegheremo, la selezione naturale è in grado solamente di selezionare variazioni dagli effetti estremi, come ciò che causa o previene la morte.
Tale principio fu riconosciuto dal famoso scienziato Motoo Kimura, che constatò che quasi tutte le mutazioni avessero un effetto troppo piccolo sull’organismo perché esse potessero essere individuate dalla selezione naturale. Egli elaborò un grafico in cui mostrava come moltissime mutazioni fossero invisibili alla selezione.

Nella parte grigia, chiamata “scatola di Kimura”, ricadono infatti tutte le mutazioni non selezionabili. Dalla sinistra dello zero sono rappresentate tutte le mutazioni che hanno un effetto negativo, mentre dalla destra dello zero sono rappresentate tutte le mutazioni dall’effetto positivo.
Ma come è facilmente capibile, tutte le mutazioni positive hanno un effetto troppo piccolo per essere selezionate, poiché ricadono nella scatola di Kimura.
Un altro evoluzionista, J.B.S. Haldane, ha determinato negli anni ’50 che i tratti necessitavano di un vantaggio di selezione di almeno 10% per far sì che la selezione naturale potesse avere una possibilità ragionevole per renderli stabili nella popolazione.
Un 10% di vantaggio di selezione significa che gli individui con questo tratto producono 10 % in più di prole che sopravvive, rispetto a quei individui senza quel tratto (Haldane, The cost of natural selection, J. Genetics 55:511-524).
Gli evoluzionisti presuppongono che ci sia circa una mutazione per individuo per generazione. Infatti, avendo riconosciuto l’effetto distruttivo delle mutazioni, hanno dovuto ipotizzare che ce ne fossero poche poiché, se non fosse così, la selezione naturale non sarebbe in grado di scartarle, e si verificherebbe il cosiddetto “genetic meltdown”, ovvero l’accumulazione di mutazioni in una popolazione che porta all’estinzione. Però negli ultimi anni è stato constatato che il tasso di mutazione è ben 50 volte più alto di quello che gli evoluzionisti avevano assunto, come dimostrato da John Sanford (già professore di genetica dell’Università di Cornell e inventore della biolistica) nel suo libro Genetic Entropy.
Oltre al tentativo di abbassare il tasso delle mutazioni, gli evoluzionisti, da ormai tanto tempo, hanno cercato di sostenere che il 97% del DNA non fosse funzionale (detto “DNA spazzatura”). In tal modo l’evoluzionista cerca di limitare la distruttività dell’assioma primario (il meccanismo mutazione + selezione naturale), ma come ha ormai provato definitivamente il progetto ENCODE (anche in questo caso il DNA spazzatura sarà oggetto di approfondimento dei nostri futuri articoli), il DNA spazzatura è un’altra speculazione darwinista, ormai abbandonata dagli evoluzionisti più onesti.
La scoperta di John Sanford crea un’enorme problema per la teoria dell’evoluzione, infatti l’alto tasso di mutazione, l’effetto distruttivo di quest’ultimo, e il fatto che le mutazioni negative che ricadono nella scatola di Kimura non possano essere selezionate (quindi né preservate e né scartate) portano ud un graduale accumulamento nel tempo di mutazioni negative in una data popolazione. Questo processo di selezione naturale e mutazione, che Sanford chiama “assioma primario”, sta portando tutti gli animali alla distruzione e all’estinzione, non al miglioramento e alla complessificazione.
L’altra implicazione che Sanford individua è che anche le mutazioni positive necessitate dall’evoluzione ricadono tutte nella zona neutrale (altro termine per “scatola di Kimura”) senza poter essere selezionate. Sanford, nel suo libro, dimostra come i singoli nucleotidi (le lettere del DNA che costituiscono l’informazione genetica, l’equivalente delle lettere che formano le parole) diano un contributo infinitesimamente piccolo all’intero genoma. La domanda è quindi “come hanno fatto ad arrivare qui e come hanno fatto a essere preservati dalla selezione?” La selezione naturale infatti è cieca, non è capace di selezionare ciò che non porta un significativo vantaggio.
Poiché un singolo nucleotide non dà quasi nessun vantaggio all’organismo, l’unico modo per essere selezionato sarebbe che in anticipo la selezione naturale preservasse quel nucleotide sapendo che, anche se non serve momentaneamente, servirà in futuro quando avverranno altre specifiche mutazioni. Ma questo è chiaramente impossibile. La gradualità che necessita il processo evoluzionistico rende impossibile la selezionabilità dei nucleotidi che servono per creare nuova informazione. Il piccolo contributo di ogni nucleotide è quindi coperto dal “rumore” di fondo di tutti gli altri nucleotidi (termine che Sanford usa e che verrà sviluppato in futuro).
Per riassumere, l’argomento di Sanford contro l’evoluzione si può sintetizzare così:
- Le mutazioni compaiono più velocemente di quanto la selezione naturale possa eliminarle.
- Le mutazioni sono troppo “modeste” nel loro effetto per essere selezionate (tranne quelle estremamente distruttive).
- La selezione naturale diventa impotente d’innanzi al rumore biologico e alla “sopravvivenza del più fortunato (non il più forte)”.
- Le mutazioni positive sono quasi sempre fisiologicamente collegate a enormi blocchi di mutazioni negative, in modo da non poter essere separate nella discendenza (per sbarazzarsi del negativo e mantenere il positivo). Il risultato è che tutti i genomi complessi stanno chiaramente degenerando. Approfondiamo un attimo il 4 punto: Detto questo, sembra evidente che il meccanismo evoluzionistico non sta facendo altro che degradare pian piano ogni organismo, ma dato lo scambio di geni provvisto dalla madre e dal padre, non potrebbe questo provvedere ad evitare la trasmissione di geni difettivi? Ovvero, non potrebbero gli spermatozoi e le uova “cattive” portare a una prole difettosa destinata a non sopravvivere, semplicemente lasciando spazio alle versioni “buone” nella popolazione? Apparentemente così si risolverebbe la questione: gli organismi difettivi vengono eliminati dalla selezione naturale, mentre quelli “sani” no. Ma purtroppo non è così, come dice il punto quattro, le mutazioni positive sono fisicamente collegate a quelle negative, senza poter essere separate. Ecco il perché: Un numero enorme di mutazioni, accumulatesi generazione dopo generazione, sono aggiunte alla linea germinale (materiale genetico che può essere trasmesso ad un discendente) di ogni neonato, e queste (mutazioni) sono diffuse in tutti i cromosomi. I nucleotidi esistono in grandi gruppi o blocchi connessi, variando in dimensione da 10,000 a un milione, ereditati in toto, senza mai spezzarsi (p 55 e 81 del libro Genetic Entropy). Pensate a questi blocchi inseparabili di nucleotidi come delle pagine. Se vuoi eliminare una mutazione (una “lettera”) da quella pagina, devi eliminare tutta la pagina. Di conseguenza, un tratto desiderabile sarà accompagnato da più tratti indesiderati, all’interno dello stesso individuo (p. 79).E’ bene sottolineare che la ricombinazione genetica non può essere usata come soluzione a questo problema poiché questa appare essere primariamente tra geni piuttosto che, in modo casuale, tra nucleotidi.Per concludere, tutti i genomi, dai più semplici ai più complessi, si stanno distruggendo ed estinguendo. Proprio l’opposto di quello che l’evoluzione necessita. La selezione naturale non è quindi in grado di fare ciò che il darwinismo necessita: prendere il buono e scartare il cattivo. Quindi, all’interno di ogni blocco di nucleotidi non separabile, di media, migliaia di mutazioni deleterie si accumulerebbero prima che una mutazione benefica compaia (p.82). Tutti i blocchi di nucleotidi inseparabili nei genomi si stanno quindi deteriorando, senza che la selezione naturale possa fare nulla.
Gli evoluzionisti informati sono consapevoli di questi problemi e come risposta hanno proposto l’epistasi sinergica, in cui gli effetti delle multiple mutazioni che accadono tutte insieme creano un effetto più grande delle loro somme. Sanford però ha dimostrato che questo renderebbe il problema peggiore. La scienziata Tomoko Ohta, nota come una delle più grandi genetiste di sempre, ha ammesso che la epistasi sinergica non fa altro che aggravare la cosa. (Sanford, Critics ignores reality of Genetic Entropy, March 2013). Inoltre, Sanford per mettere definitivamente a tacere questa opzione, fece delle simulazioni matematiche che usano un modello di popolazione genetica chiamato “Mendel’s Accountant”, dimostrando ancora una volta le fragilità delle obiezioni evoluzionistiche.
Per concludere, non si può credere che un processo che non fa altro che distruggere genomi possa creare un organismo migliore.
BREVE ILLUSTRAZIONE DEL PROBLEMA EPIGENETICO INDIVIDUATO DA STEPHEN C.MEYER
Nel libro di Stephen C. Meyer, Darwin’s Doubt, si indaga sul dubbio centrale che Darwin ebbe della sua stessa teoria, cioè che in tutti i reperti fossili non vi erano le forme intermedie che la sua teoria di cambiamenti evoluzionistici graduali richiedeva. Sebbene preoccupato per la mancanza dei fossili transizionali, Darwin non ne fece un grande problema, sperando che le scoperte future avrebbero confermato la sua teoria. Meyer sostiene che queste scoperte non sono mai avvenute. Infatti qualsiasi ricerca approfondita tra i reperti fossili e conferma oggi ciò che originariamente Darwin stesso notò: la comparsa discontinua e brusca delle prime forme di vita animale complessa in un remoto periodo geologico della storia conosciuto come il periodo Cambriano. Infatti, i paleontologi ora ritengono che approssimativamente venti dei ventisei phylum animali (che rappresentano i distinti piani corporali) trovati nell’insieme dei reperti fossili appaiono senza predecessori in un evento geologico senza paragoni chiamato “l’esplosione Cambriana”.
Di certo Darwin non era a conoscenza del DNA o dell’informazione biologica aggiuntiva necessaria per rendere possibile la vita. Se avesse conosciuto questi fatti scientifici, avrebbe capito che i reperti fossili non erano l’unico problema della sua teoria. Adesso sappiamo che i nuovi piani corporali, come quelli che osserviamo essere comparsi improvvisamente nell’era Cambriana, necessitano di milioni di nuovi caratteri di precisa e codificata informazione genetica, e che tutta questa nuova informazione doveva essere nata velocemente per spiegare l’improvvisa comparsa. Anche se accettiamo per ipotesi che la prima forma di vita avesse già informazione genetica preesistente, come è possibile che cieche forze della natura possano creare nuova informazione per nuovi piani corporali (per farla molto semplice sono un gruppo di caratteristiche strutturali e di sviluppo che può essere utilizzato per identificare un gruppo di animali, come il phylum)? Una mutazione di informazione preesistente non funzionerebbe poiché tali mutazioni sono quasi sempre dannose. Quanto tempo un lettore impiegherebbe a distruggere il senso di questa stessa frase spostando casualmente queste lettere? Quanto tempo Microsoft Word potrebbe funzionare se il suo codice incominciasse a mutare casualmente? Smetterebbe molto rapidamente di funzionare (il nostro DNA è un insieme di lettere strutturate grazie ad un codice ed una sintassi. Ne parleremo nell’Argument from information).
Ma questo è un problema che abbiamo già trattato nella parte 1.
La premessa appena fatta serve ad introdurre brevemente un ulteriore problema individuato da Stephen C. Meyer nel 14 capitolo del suo libro Darwin’s Doubt.
Possiamo definire questo problema come il “problema epigenetico”: anche se ci fosse tempo infinito e opportunità infinite affinché la natura possa mutare il DNA in quella informazione necessaria per nuova vita, questo ancora non basterebbe per creare una nuova forma di vita, questo perché il DNA da solo non detta la formazione di piani corporali.
Negli ultimi anni i biologi hanno scoperto una nuova forma di informazione essenziale (necessaria) per la formazione del corpo chiamata informazione epigenetica, la quale non è contenuta nel DNA ma nelle strutture della cellula. L’informazione epigenetica è quella che è impartita dalla forma e dalla struttura della cellula embrionale includendo l’informazione dell’uovo. In parole semplici, la struttura fisica delle cellule ancora ad uno stadio iniziale traccia un percorso di sviluppo per l’organismo.
Nel 14 capitolo di Darwin’s Doubt si descrivono i diversi tipi di informazione epigenetica che i biologi hanno individuato. I dettagli verranno analizzati in futuro, ma in breve si può dire che costruire un essere vivente necessita:
1) Il DNA per fare le proteine
2) Le proteine devono essere organizzate in strutture cellulari e i diversi tipi di cellula
3) I tipi di cellula devono poi essere organizzati in tessuti
4) I tessuti devono poi essere organizzati negli organi
5) Gli organi e tessuti devono poi essere organizzati nei piani corporali
Anche se il DNA contribuisce in tutti e 5 i passi, l’informazione epigenetica è necessaria per spiegare i passi 2,3,4,5. Il DNA non è infatti in grado di farlo da solo.
Pensate a questo esempio: una casa da costruire. Per far sì che si possa costruire una casa, si ha bisogno di specifiche istruzioni e i materiali fisici di una certa dimensione e composizione, come viti, chiodi, cemento e fili elettrici, ed infine questi materiali devono poi essere posizionati in una struttura precisa. Allo stesso modo, per costruire un essere vivente, sono necessitate specifiche istruzioni (il DNA), e i materiali fisici di una certa misura e composizione, che devono essere formate in specifiche strutture come le cellule, i tessuti, gli organi… (informazione epigenetica). Il DNA può solo spiegare quindi l’esistenza dei specifici materiali, e non della loro organizzazione in piani corporali.
Detto questo, non è possibile costruire una casa più complessa di quella precedente semplicemente cambiando il libretto di istruzioni della tua casa. Allo stesso modo non si può costruire una forma di vita più complessa semplicemente mutando le istruzioni scritte di una forma di vita esistente. Nuovi materiali e strutture sono necessitate.
Molto semplicemente, si può dire che l’informazione epigenetica orchestri dall’alto il tutto, mentre il DNA spiega la formazione dei singoli pezzi, o meglio le proteine, ma non è in grado di farle funzionare assieme.
Ma non può essere mutata l’informazione epigenetica per creare nuove forme di vita? Meyer sostiene che questa sia una delle domande più comuni che egli riceve e la sua risposta e sempre “no” per diversi motivi, il principale è che un cambio epigenetico ucciderebbe l’organismo immediatamente.
L’implicazione di questo argomento è di fondamentale importanza: infatti poiché il DNA da solo non è in grado di controllare la formazione dei piani corporei, semplicemente mutare il DNA non creerà mai un nuovo piano corporeo. La teoria dell’evoluzione è quindi falsa.
Questo argomento non si basa nemmeno sulla probabilità: anche se il tempo a disposizione fosse infinito, l‘organismo non avrà mai un piano corporeo diverso dal suo. Meyer dice: “Anche nel migliore dei casi, per uno che ignora l’immensa improbabilità della generazione di nuovi geni tramite mutazioni e selezione naturale, le mutazioni nella sequenza del DNA creerebbero semplicemente nuova informazione genetica. Ma costruire un nuovo piano corporeo richiede più che solamente informazione genetica. Richiede informazione sia genetica che epigenetica, informazione che per definizione non è immagazzinata nel DNA e quindi non può essere generata da mutazioni del DNA. Ne consegue che il meccanismo della selezione naturale che opera su mutazioni casuali nel DNA non può da solo generare nuovi piani corporei, come quelli che comparvero nell’esplosione cambriana” (Stealing from God, Frank Turek, 63).
L’unico sostanziale tentativo di confutazione di questo argomento proviene dal rispettabile paleontologo Dr. Charles Marshall, che recensì “Darwin’s Doubt” sulla prestigiosa rivista Science. Il Dr. Marshall asserì che nuovi piani corporali nacquero attraverso il “ricablaggio” (riorganizzazione) di geni già esistenti (o meglio, di network genetici regolatori).
Meyer rispose che anche se ciò fosse vero, il “ricablaggio” di geni esistenti richiederebbe esso stesso un’infusione di un codice genetico aggiuntivo. Inoltre, sappiamo dagli esperimenti che (rimescolando) le reti di regolazione genetica, che sono sistemi di controllo per il funzionamento della cellula, si causa inevitabilmente la morte dell’organismo.
La posizione di Marshall non fornisce inoltre alcuna spiegazione naturalistica per l’origine dei geni stessi; semplicemente presuppone la loro esistenza. Dunque Marshall non ha risolto il problema dell’orine della informazione biologica, l’ha semplicemente spostato più in là.
Quando il Dr. Meyer ha insistito su questa risposta in un dialogo via radio, il Dr.Marshall ha semplicemente risposto: “buona osservazione”! (Unbelievable?, Darwin’s Doubt – Stephen C. Meyer & Charles Marshall debate ID – Does the Cambrian explosion support Intelligent Design? November 29, 2013).
Altri due problemi statistici
Oltre al problema statistico di Behe, il quale asserisce che funzioni che necessitano di più di due mutazioni all’interno del genoma non hanno il tempo materiale per formarsi, esistono molti altri problemi statistici.
1) IL DILEMMA DI HALDANE
L’evoluzionista J.BS. Haldane fu uno dei fondatori della genetica delle popolazioni e riconobbe seri problemi nella teoria darwinista. I suoi studi infatti dimostrarono che il processo di selezione naturale è troppo lento per poter forare la vita oggi presente.
Quando una mutazione benefica nasce in una popolazione, deve incrementare in frequenza per far sì che la popolazione evolva (se la mutazione rimanesse in un solo individuo, allora l’evoluzione non potrebbe procedere, questo è ovvio). In altre parole, tale mutazione deve sostituirsi ai geni non mutati all’interno della popolazione. Ma il tasso con cui questo può accadere è ovviamente limitato.
Ad esempio, uno dei principali fattori che limita il tasso di sostituzione è il tasso di riproduzione delle specie. Ad esempio, il tasso di generazione dell’essere umano è di circa 20 anni e ha un tasso di riproduzione per individuo abbastanza basso, quindi il tasso di crescita di un mutazione in una popolazione sarà estremamente basso.
Immaginate una popolazione di 100,000 scimmie, i supposti progenitori dell’uomo. Immaginate adesso che sia in una scimmia femmina sia in una scimmia maschio si riscontri una mutazione così benefica da farli rimanere gli unici esemplari a sopravvivere: infatti tutte le altre 99,998 scimmie morirono. E la coppia sopravvissuta ebbe abbastanza tempo per riprodursi per ricostituire numericamente la popolazione in una generazione.
Immaginate adesso che questo succeda ad ogni generazione (ogni 20 anni) per 10 milioni di anni, più del tempo che gli evoluzionisti dicono essere passato dall’ultimo antenato comune tra la scimmia e l’uomo. Questo vorrebbe dire che 500,000 (ovvero 10 milioni diviso 20 milioni) mutazioni benefiche si sarebbero aggiunte alla popolazione.
Anche con questo scenario completamente non realistico, che massimizza alle stelle le condizioni perfette per un processo evolutivo, si formerebbe solo circa il 0,02% del genoma umano!
Considerando che il DNA di un essere umano e quello di una scimmia, il nostro supposto parente più vicino, hanno almeno 150 milioni di basi (lettere) diverse, ovvero non meno del 5% di differenza. (Tomkins, Bergman, Genomic Monkey Business, J. Creation 26 (1): 94-100, 2012)
Queste considerazioni mostrano che la teoria evoluzionistica si trova davanti ad un enorme problema, in quanto è stato mostrato come la fissazione nella popolazione di un nuovo tratto impieghi un tempo enorme.
E’ proprio per questo, e altri motivi, che gli evoluzionisti inventarono il concetto di DNA spazzatura. Gli evoluzionisti realizzarono infatti che la selezione naturale non poteva formare abbastanza DNA (appunto solo una piccola percentuale di quello che è il nostro DNA), allora ipotizzarono che la maggior parte del DNA non fosse funzionante.
Tenendo conto dei tassi più realistici di adattamento, selezione e tasso di riproduzione, il numero di mutazioni benefiche che può essere accomodato/inserito in una popolazione crolla. Haldane calcolò che non più di 1,667 sostituzioni benefiche possono essere avvenute nei supposti 10 milioni di anni (per essere generosi) dall’ultimo antenato dell’uomo e della scimmia. Ciò significa che può verificarsi circa una sostituzione ogni 300 generazioni.
Haldane intese il termine sostituzione come un singolo evento di mutazione che si diffonde in tutta la popolazione (si fissa). Tale mutazione può quindi verificarsi in una duplicazione genetica, un’inversione cromosomica o una sostituzione di un singolo nucleotide, un’inserzione o un’inversione.
I biologi hanno trovato che la grande maggioranza delle mutazioni sono in effetti sostituzioni di singoli nucleotidi, dunque il limite di Haldane pone una severa costrizione riguardo a ciò che è possibile ottenere con il processo evoluzionistico, perché 1667 singole sostituzioni di nucleotidi ammontano a meno di un gene (di grandezza media)!
Va inoltre notato che Haldane fece delle presupposizioni semplificatorie che invece ottimizzarono il numero di sostituzioni che sarebbero possibili. Dunque in realtà il numero di sostituzioni possibili è molto minore di quello che ha calcolato Haldane.
L’analisi di Haldane mostra che la selezione naturale di mutazioni non può essere responsabile per i genomi degli organismi come gli uomini (o per altri casi specifici, come ad esempio la formazione del collo delle giraffe), in quanto sarebbe di gran lunga troppo lenta. Gli evoluzionisti hanno sempre tentato di evitare questo problema, ma recentemente Walter ReMine ha affrontato il problema con una analisi che cerca anche di chiarire la confusione che ebbe Haldane sull’argomento. (Batten, D., Haldane’s dilemma has not been solved, J. Creation 19(1):20-21, 2005; creation.com/haldane)
2) L’IMPOSSIBILITA’ STATISTICA DELLA FORMAZIONE DI PROTEINE DIMOSTRATA DALLO SCIENZIATO DOUGLAS AXE
Dato che i geni (porzioni del genoma) codificano le proteine, e nuove proteine sono necessarie per una evoluzione darwinista, Douglas Axe studiò la probabilità della formazione di nuovi geni capaci di codificare nuove proteine.
Prima di iniziare a descrivere il suo studio è bene sottolineare come i geni capaci di produrre nuovi “ripiegamenti di proteine” hanno solo due modi per formarsi: a) o da geni preesistenti o b) da sezioni non funzionanti del genoma.
Per “ripiegamento di proteina” si intende la forma/configurazione funzionale delle proteine (si veda immagine: a sinistra c’è la catena di amminoacidi e a destra essa assume un particolare forma che le conferisce una particolare funzione diventando proteina).

Nel primo caso (a), gli evoluzionisti potrebbero sostenere che gradualmente un gene che codifica una proteina muti fino a riuscire a codificare un’altra proteina.
Sostenere questo primo scenario è però irragionevole e ciò è riconosciuto praticamente da tutti gli evoluzionisti. I motivi per il quale la prima opzione è considerata ormai da rigettare sono i seguenti:
Mutazioni in geni preesistenti rovinerebbero e degraderebbero l’informazione funzionale di quei geni. Quando un gene perde la sua funzione (codificare le proteine) la selezione eliminerà l’organismo che possiede questo gene. Questo processo si chiama “selezione purificatrice”.
Douglas Axe nel suo esperimento “Extreme Functional Sensitivity to Conservative Amino Acid Changes on Enzyme Exeriors” dimostra che è incredibilmente difficile provocare dei cambiamenti ad una sequenza amminoacidica (catena di amminoacidi nell’immagine a sinistra) funzionale, senza destabilizzare il ripiegamento delle proteine. Trovò che come alterava questa proteina, diversi cambiamenti di posizione all’esterno della molecola di proteina facevano sì che la proteina perdesse la sua funzione.
Infatti, per passare da una proteina con un certo ripiegamento ad una proteina con un ripiegamento e con una struttura e funzione completamente nuova, si richiedono specifici cambiamenti in tanti, tantissimi punti. Infatti il numero di cambiamenti necessari per produrre un nuovo ripiegamento di proteina eccede il numero di cambiamenti necessari che causerebbero la perdita della sua funzione.
Detto questo, per fare un’analogia, è come se il processo evoluzionistico dovesse fare un salto da una cima di una montagna all’altra, senza passare per la valle. La cima rappresenta la proteina funzionale e la valle il grande spazio di condizioni senza alcuna funzionalità, ovvero proteine non funzionali o catene amminoacidiche non in grado di ripiegarsi.
L’incapacità del processo evoluzionistico è direttamente proporzionale al numero di cambiamenti necessitati per passare ad un ripiegamento di proteina ad un altro. Il lavoro di Axe sopracitato dimostra l’altissima sensibilità delle proteine al cambiamento, con il suo esperimento infatti è stato dimostrato che la perdita di funzione è la conseguenza immediata del cambiamento.
C’è un ulteriore motivo per credere che l’evoluzione di proteina in proteina sia irragionevole: quasi tutte le funzioni delle proteine non possono essere eseguite se le proteine non sono ripiegate. Destabilizzare quindi, anche con il più lieve cambiamento, i ripiegamenti di proteine fa sì che la proteina perda la sua struttura tridimensionale perdendo la sua funzione.
Per di più le proteine destabilizzate vengono attaccate da altre proteine, le proteasi, che letteralmente distruggono le proteine non ripiegate e i polipeptidi nella cellula!
Detto questo, se anche ciò che ha una piccola perdita di funzione viene eliminata dalla selezione naturale, figuriamoci ciò che perde totalmente la sua funzione.
Axe concluse quindi che, anche con il mimino cambiamento la proteina si sarebbe distrutta o avrebbe perso la sua funzionalità.
Tutte queste affermazioni sopracitate sono poi state confermate numerose volte per via sperimentale, come nel caso degli studi del famoso biologo molecolare Francisco Blanco, nel suo “Exploring the Conformational Properties of the Sequence Space Between Two Proteins with Different Folds”
La prima opzione di evoluzione dei geni è quindi sbagliata.
Per questi motivi Axe si concentrò sullo studio dell’opzione apparentemente più plausibile, ovvero la seconda (b), in cui nuovi geni si formano da regione non funzionali o neutre del genoma.
In questo scenario l’informazione che compare da sezioni neutre o non funzionali del genoma, che possono essere quelle che non codificano per le proteine, o quelle duplicate, può variare liberamente, senza danneggiare l’organismo. Infatti, alterare i nucleotidi del DNA in queste zone non provoca nessun effetto sull’organismo, ed è per questo che la selezione naturale non ha nessun ruolo, almeno finché un nuovo gene funzionale non è comparso.
Gli evoluzionisti, per il più delle volte, sostengono che questo processo di formazione dei geni derivi da una duplicazione avvenuta durante il processo del crossing-over nella meiosi (un tipo di divisione cellulare che produce gameti in organismi sessuati).
Una volta avvenuta la duplicazione, che in parole povere sta a significare una copiatura di un gene già esistente, si permette ad uno dei due geni di poter essere modificato da varie mutazioni, senza che l’organismo ne subisca le conseguenze deleterie, poiché l’alto gene continua a funzionare normalmente. La conseguenza di tale modificazione è che è selettivamente neutra, poiché, dato che non è né in grado di creare danni e né in grado di creare benefici (ameno fin quando non si forma un nuovo gene con una nuova funzione). Diviene quindi possibile sperimentare e variare liberamente in sicurezza, facendo sì che questi nuovi cambiamenti genetici ininfluenti possano essere passate alle generazioni successive, dando così la possibilità in futuro ad altre mutazioni di far continuare ad “evolvere” questo gene, creando una nuova sequenza capaci di codificare per una nuova proteina. Questo secondo modello è definito dagli evoluzionisti il “modello classico” dei evoluzione di un gene.
Se da una parte questo modello è superiore al primo in quanto, la transizione del gene originale al nuovo gene non comporta alcun effetto distruttivo, tuttavia incontra un problema sostanziale, ed è proprio questa l’argomentazione di Axe: le sequenze capaci di formare ripiegamenti di proteine che rimangono stabili e che svolgono una funzione biologica sono estremamente poche (rispetto al numero totale di sequenze possibili). Poiché la selezione naturale non può fare nulla per mantenere una sequenza transizionale, ovvero una catena di amminoacidi che è a “metà strada” tra la proteina che era originariamente e quella che deve diventare, tramite graduali cambiamenti di posizione degli amminoacidi stessi, è solamente compito delle mutazioni creare una sequenza estremamente rara, capace di codificare per un ripiegamento funzionale (la selezione naturale non può prevedere che un codice apparentemente inutile possa servire in un ipotetico futuro),.
L’esperimento “Esperimenting the Prevalence of Protein Sequences Adopting Functional Enzyme Folds” di Axe consistette, tramite la “mutagenesi sito diretta”, di studiare il rapporto della frequenza tra le sequenze capaci di codificare per un ripiegamento funzionale di proteina ed una sequenza inutile (ovvero non capace di codificare per una proteina funzionale).
Egli stimò, con altissima precisione, che una sequenza di 150 amminoacidi capace di ripiegarsi in una struttura tridimensionale (una proteina appunto), comparata all’intera possibilità di sequenze amminoacidiche di quella lunghezza ha un rapporto di 1 su 1074.In altre parole, esiste una sola sequenza di amminoacidi capace di ripiegare una proteina per ogni 1074 sequenze.
Poiché una proteina per funzionare deve essere ripiegata, ma non tutte le proteine ripiegate sono funzionali, Axe calcolò che le sequenze capaci di produrre un ripiegamento di una proteina che abbia una funzione siano 1 su 1077.
Pensate ad esempio ad un qualsiasi sistema linguistico: per una parola di 10 lettere esistono molte più sequenze di lettere che non creano alcun significato rispetto a quelle che creano una parola con significato, tenendo a mente che il DNA, è composto da nucleotidi, come questo articolo è composto da lettere, cambiando la posizione dei nucleotidi, si può cambiare il messaggio del DNA. In questo caso, il gene, che è l’insieme di nucleotidi che codifica per una proteina, dice esattamente in che posizione devono collocarsi i precisi amminoacidi per far sì che si formi una proteina funzionale. Cambiando quindi la sequenza nucleotidica si può cambiare la sequenza amminoacidica per creare nuove forme di proteine necessarie per un processo evoluzionistico. Questa sequenza di 150 amminoacidi per diventare funzionale, tramite mutazioni, in modo del tutto cieco (la selezione naturale non può aiutare), deve azzeccare l’esatta sequenza che permetta il ripiegamento. Ricordiamoci però che solo una sequenza su 1077crea un ripiegamento funzionale! Tenete a mente che la somma di tutti gli atomi della Via Lattea ammonta a 1065!
Detto questo, non rimane altro che valutare la probabilità di questo evento (della nascita di una sequenza funzionale) dato il numero di tentativi stimati.
Il tempo gioca qui un ruolo fondamentale. Prendete ad esempio l’analogia del ladro che cerca la combinazione del lucchetto la bicicletta è legata. Trovare la combinazione di un lucchetto che necessita l’inserimento di 4 cifre, è difficile ma possibile se al ladro è dato qualche giorno per provare tutte e combinazioni possibili. Infatti ogni lucchetto di 4 cifre ha 10,000 tipi diversi di combinazioni di cui solo una apre il lucchetto. Il problema sussiste quando il ladro ha un tempo a disposizione molto limitato, supponiamo un’ora. A questo punto diventa improbabile che trovi la combinazione. E qui che entra in gioco la probabilità condizionale, ovvero, in parole semplici, la probabilità che tiene conto di diversi fattori, come appunto il tempo.
Axe, era interessato al numero di mutazioni, che nel corso della storia della vita sulla Terra, avrebbero potuto creare una nuova sequenza di basi nel DNA capace di creare una nuova sequenza di amminoacidi, una delle 1077 sequenze possibili. Infatti maggiori sono i tentativi e più aumentala probabilità della formazione di una proteina. Eppure bisogna tenere conto che non tutte le mutazioni possono essere considerate tentativi validi per la ricerca della sequenza giusta. Possono essere considerati tentativi validi infatti solo le mutazioni che possono essere ereditate. Quindi possiamo solo tenere conto delle mutazioni dei geni che avvengono nelle cellule riproduttive.
Detto questo, si può fare una stima del numero totale di organismi che sono vissuti sulla storia della Terra e il numero di geni che le mutazioni potrebbero produrre e trasmettere alla prole. Tenendo conto delle enormi popolazioni di procarioti, come i batteri, e assumendo che la vita sia nata sulla Terra 3.8 miliardi di anni fa, Douglas Axe (e numerosi altri scienziati) ha stimato che dalla prima cellula sono vissuti sino ad oggi 1040 animali. Axe, inoltre, presuppose che ogni organismo ricevette una nuova sequenza di basi, ovvero potenzialmente un nuovo gene, capace di produrre una delle possibili sequenze amminoacidiche per generazione.
Questa presupposizione è incredibilmente a favore degli evoluzionisti. Poiché le mutazioni devono essere abbastanza rare per far sì che l’organismo possa sopravvivere, tantissimi batteri ereditano una copia esatta di DNA dai loro genitori. Inoltre, quando viene trasmesso il DNA con una mutazione è molto probabile che quella mutazione sia già avvenuta molte volte in altri batteri. Per questo motivo il numero attuale di sequenze tentate a partire dalla prima cellula, è molto inferiore del numero totale di tutti i batteri mai esistiti sulla Terra.
In ogni caso, assumendo che un nuovo gene per organismo sia stato trasmesso alla generazione successiva, Axe concluse che dall’inizio della prima vita ci sono stati 1040 tentativi alla ricerca della formazione di una sequenza che creasse una nuova proteina. Quindi la probabilità condizionale della formazione di un nuovo gene capace di codificare per un nuovo ripiegamento di proteina è una su 1037 (1077– 1040).
Tanto per complicare le cose sottolineiamo che: primo, l’esplosione cambriana datata mediante i ritrovamenti fossili 600 milioni di anni fa (per essere generosi), riduce di gran lunga il tempo per la formazione degli animali. Meno tempo equivale a dire meno tentativi, e quindi maggiore improbabilità. Secondo, i batteri sono gli animali che più hanno costituito la stima complessiva degli animali sulla Terra, eppure nessun animale del Cambriano si è evoluto direttamente dal batterio, e nemmeno nessuno crede che i primi organismi pluricellulari fossero così abbondanti come nel caso dei batteri. Infatti un numero minore dei possibili antenati dei primi animali ridurrebbe ulteriormente la già alta improbabilità: come chiunque sostiene, gli antenati pluricellulari dei primi animali del cambriano furono di numero di gran lunga meno numerosi, rispetto all’enorme numero totale di organismi mai vissuti sulla Terra. Terzo, per far evolvere un animale, sono necessari ben più di una nuova proteina, tra l’altro di una lunghezza modesta (150 amminoacidi). Tutti gli animali infatti per evolvere necessiterebbero di proteine ben più lunghe, rispetto a quella presa in considerazione da Axe, come nel caso della proteina chiamata “lisil ossidasi”, necessaria per l’evoluzione anche dei primissimi animali del cambriano, che è composta da ben 400 amminoacidi.
Prendendo sempre in considerazione il Cambriano, gli animali comparsi in questo periodo, per evolversi dagli organismi pluricellulari, necessiterebbero tantissime cellule diverse rispetto a quelle degli antenati. Queste cellule sono composte da numerose differenti proteine, il che complica ancora di più la vita agli evoluzionisti.
Tra l’altro, l’alta improbabilità di una sequenza funzionale (1077) spiega anche come mai il primo caso dell’evoluzione dei geni (da un gene preesistente appunto) non possa avvenire.
Per concludere gli evoluzionisti devono risolvere questo dilemma: se la selezione naturale non gioca alcun ruolo, subentra il problema statistico sopracitato, se invece la selezione naturale esercita una pressione, come nel caso dell’evoluzione da un gene già esistete, si presentano i problemi sopracitati.
Il meccanismo evolutivo non è quindi in grado di creare informazione genetica.
Esistono molti altri problemi statistici contro la teoria dell’evoluzione, che nel tempo analizzeremo singolarmente.
LA COMPLESSITA’ IRRIDUCIBILE
La complessità irriducibile è considerata da molti l’argomento più efficace contro la teoria dell’evoluzione. L’evoluzione infatti necessita di una gradualità nei processi (tanto per intenderci, nessun evoluzionista crede sia possibile passare da un pesce ad un anfibio in una generazione) dovuta al lento meccanismo di mutazione e selezione naturale.
Studiando questi sistemi che sono appunto irriducibilmente complessi (come dice la parola stessa, non possono essere ulteriormente scomposti nelle sue parti senza perdere la loro funzione) vediamo come un processo graduale come quello darwinista non può spiegare la loro esistenza.
Infatti, la transizione diviene impossibile in un sistema in cui, anche senza solo una delle sue parti, esso non funzionerebbe, poiché, assumendo il gradualismo, le transizioni del sistema in questione sarebbero completamente inutili e non avrebbero motivo per essere preservate dalla selezione naturale (la sua funzione infatti sarebbe possibile solo con tutte le sue parti già esistenti).
Un sistema è irriducibilmente complesso quando richiede che diverse parti collegate fra loro siano esistenti allo stesso momento, mentre quando avviene una rimozione di una o più di queste parti il sistema non funzionerà. Distruggere anche solo una parte del sistema, lo farebbe crollare. Il presunto meccanismo evolutivo, d’altro canto, implica che una nuova caratteristica porterà vantaggi per la sopravvivenza, e dunque permettere all’organismo di competere meglio contro organismo che non hanno “acquisito” questo tratto.
La teoria evoluzionistica necessita che tutte le forme di vita possano essere riconducibili a forme di vita sempre più semplici fino ad arrivare alla cellula primordiale, che a sua volta è venuta dalla materia, cosa ancora più semplice. Darwin, relativamente a questo disse: “Se fosse dimostrato che esiste un organo complesso che non poteva essere stato formato da numerose, successive, lievi modificazioni, la mia teoria si sgretolerebbe.” (Charles Darwin, L’origine delle Specie, 1964, p.189).
Michael Behe, nel suo libro “La scatola nera di Darwin”, illustra in modo dettagliato diversi sistemi da lui considerati irriducibilmente complessi. In questo articolo tratteremo solo uno di questi casi in modo molto riassuntivo. In futuro descriveremo dettagliatamente questi sistemi, con le critiche sollevate e le relative confutazioni delle critiche. Altri sistemi irriducibilmente complessi che tratteremo successivamente sono ad esempio tra i moltissimi, la coagulazione del sangue, il sistema immunitario, il motore ATP, il motore Kinesin, il sistema di trascrizione del DNA, la cellula stessa (nel tempo analizzeremo anche altri casi di complessità irriducibile).
Un esempio di complessità irriducibile: il flagello del batterio
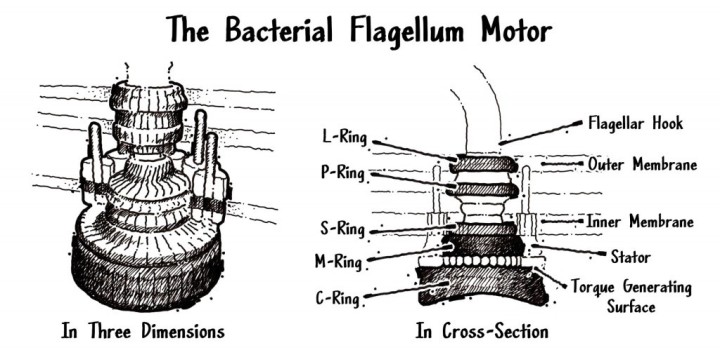

Alcuni batteri nuotano ruotando un lungo filamento che può ricordare una frusta. Questo motore rotante assemblato è chiamato flagello. I flagelli dei batteri sono incredibilmente difficili da spiegare per gli scienziati che li riconoscono come una meraviglia della precisione meccanica. Il biofisico di Harvard Howard Berg ha pubblicamente descritto il flagello dei batteri come “il macchinario più efficiente dell’Universo”.
Il flagello è costituito da ben 40 parti interconnesse, e se si toglie solo una di queste parti, l’intero motore perde la sua funzionalità.
Tentando di rifiutare la potente critica all’evoluzionismo, dovuta alla complessità irriducibile del flagello, alcuni hanno proposto una “scorciatoia”. Il filosofo Robert T. Pennock crede che il complesso flagello si sia potuto formare attraverso un processo evolutivo per il quale un micro-macchinario meno complesso è preso dall’interno della cellula e utilizzato per creare qualcosa di nuovo.
In generale questa fragile obiezione alla complessità irriducibile si chiama “co-option”, in quanto si cerca di spiegare come una forma intermedia possa servire a qualcos’altro. Alcuni credono quindi che macchinari irriducibili non siano irriducibili un quanto parti transizionali di questi macchinari possono comunque essere utili, in modo diverso, all’organismo.
Alcuni scienziati hanno proposto il Sistema di Secrezione di Tipo III (T3SS, guardare immagine sopra) come esempio perfetto di uno di questi micro macchinari “prestati” per passare rendere possibile la transizione.
I T3SS sono sonde sensoriali a forma di ago usate dai batteri; individuano la presenza di organismi che possono infettare il batterio e su di essi secernono proteine per combattere il processo infettivo. I T3SS condividono molte proteine in comune e sono costruiti in modo simile al flagello di un batterio.
Gli scienziati a volte con questi T3SS tentano di spiegare come il processo evolutivo possa saltare la divisione da una singola proteina per arrivare alla complessità di un flagello. Utilizzando i T3SS, i flagelli trovano un significativo punto di partenza su cui edificarsi, diminuendo il gap di complessità. Questo approccio è però problematico per tre importanti motivi:
1) Il micro macchinario utilizzato è anch’esso irriducibilmente complesso
I T3SS sono sorprendentemente irriducibili, proprio come i flagelli.
Il T3SS è costruito da circa 30 differenti proteine ed è uno dei sistemi di secrezione più complessi in assoluto osservati oggi in biologia. Come il flagello, il T3SS richiede la configurazione basica di queste proteine per funzionare. Non può essere infatti presentato come spiegazione ultima per la complessità irriducibile poiché la sua stessa complessità irriducibile richiede una spiegazione.
Il famoso scienziato anti evoluzionista William Dembski si esprime in merito così: “Ciò che abbiamo qui non si tratta di una via pienamente articolata, ma di un’isola (il Tipo III sistemi di secrezione) e di un enorme salto per giungere alla prossima isola (il flagello). Se l’evoluzionismo tentasse di spiegare come è possibile fare un balzo da Los Angeles a Tokyo, praticamente quello che gli evoluzionisti hanno trovato sono le isole Hawaii e nient’altro. Ciò che gli evoluzionisti non hanno trovato è l’intero arcipelago [gruppo di isole collegate le une alle altre] che può portarti da una sponda all’altra.
2) Il Passaggio verso e da il micro macchinario utilizzato è empiricamente non supportato
Inoltre, come sostiene Dembski, non c’è alcuna evidenza che spieghi la graduale progressione evoluzionistica verso l’irriducibilmente complesso T3SS (da una singola proteina), e né vi è alcuna evidenza che spieghi la progressione evoluzionistica graduale dal T3SS al flagello. Infatti molti naturalisti attaccando la loro speranza nel T3SS sono però incapaci a descrivere passo dopo passo l’evoluzione da una proteina al complesso T3SS.
Il biologo H. Allen Orr dell’Università di Rochester riconosce i limiti delle spiegazioni evoluzionistiche che dipendono da questo utilizzo del T3SS: “Possiamo pensare che alcune delle parti di un sistema irriducibilmente complesso si siano evolute mano a mano per qualche altro motivo e che poi fossero reclutate in massa per una nuova funzione. Ma questo è improbabile. Si può comunque sperare che metà delle trasmissioni della tua macchina possano per caso innescare il tuo airbag nel momento del bisogno. Però tale cosa è molto, molto, molto improbabile e certamente non offre una soluzione generale alla complessità irriducibile.”
La proposta offerta dagli scienziati che cercano di spiegare il flagello con il prestito del T3SS sono affascinanti, ma prive di supporto scientifico. Quando si esaminano questi tentativi esplicativi, bisogna guardare attentamente alle costruzioni intermedie necessarie per arrivare da un micro macchinario ad una altro. Quando questi passaggi sono attentamente esaminati, essi non fanno altro che rilevare dilemmi e ostacoli.
3) Il micro-macchinario utilizzato potrebbe non essere disponibile per l’utilizzo
Gli evoluzionisti sono sempre più scettici riguardo al supposto contributo che il T3SS abbia potuto dare per costruire il flagello. Molti esperti riconoscono le similarità strutturali tra i due micro macchinari, ma rigettano ogni particolare gerarchia, ordine, collegamento evoluzionistico. Parecchi scienziati infatti ritengono che il T3SS non sia un precursore evoluzionistico del flagello, ma che sia più ragionevolmente un prodotto di una involuzione dal flagello.
Se il T3SS fosse indisponibile prima dell’esistenza del flagello, non potrebbe essere offerto come spiegazione per il flagello. Il ricercatore Jonathan Witt riassume il problema multiforme risultato: “Punto primo, la micro siringa (il T3SS) al massimo potrebbe spiegare solamente dieci proteine, lasciandone trena o più inspiegate, e queste altre trenta proteine non sono trovate in nessun altro sistema vivente. Punto secondo, come suggerisce una vasta letteratura scientifica, il sistema probabilmente si sviluppò in seguito al più complesso flagello, non al contrario. Infine, anche se la natura avesse in mano tutte le parti di proteine per costruire un flagello batterico, qualcosa dovrebbe comunque averli assemblati assieme in un preciso ordine temporale, proprio come le autovetture sono assemblate nelle fabbriche. Come è possibile questo?”.
Per un maggiore approfondimento sul T3SS leggere: “Why the Type III Secretory System Can’t Be a Precursor to the Bacteria Flagellum” di Casey Luskin.
Amedeo Da Pra e Edoardo Da Pra

3 pensieri su “INTRODUZIONE ALL’ANTI EVOLUZIONISMO – PARTE 2”